
Week 4 Blog Post – Platyhelminthes – Revised 10.08.17
Posted by kflani on September 22, 2017
Filed under Platyhelminthes
Behavior of Trematoda
Introduction
Each week of Invertebrate Zoology begins with a game. As a class we race against one another to identify unknown invertebrates that our professor presents. Upon correct identification, we choose what topic we will investigate for the week about the ~newly identified~ organism. Here’s our picture for this week. Can you guess what phylum it belongs to?

Jim Collins, Ana Vieira, and Phillip Newmark – National Geographic, 2013. Our invertebrate of the week.
Based on the organism’s soft-bodied, bilaterally symmetrical, unsegmented, and flat appearance, we guessed it was a platyhelminth – or a flatworm. We were correct!
This phylum consists of four classes: turbellaria, cestoda, monogenea, and trematoda1. Turbellaria are free-living flatworms, some of which exhibit commensalism – a relationship between two organisms where one organism benefits from the actions of the other without inflicting harm2. Cestoda, commonly known as tapeworms, are endoparasitic meaning they live within hosts (usually multiple) and eventually inflict harm or death for their own benefit1. Monogenea are typically ectoparasites (living outside) of fishes and only live depend on one host1.
Trematoda – the class of our unknown invertebrate – are endoparasites that are divided into two orders: aspidogastrea and digenea1,2,3. Aspidogastrea have simple life cycles (similar to monogenea) and usually only rely one one host – some species require an intermediate host; however, most trematodes are digenean (literally “two births”) exhibiting two life stages – miracidia and cercariae – that invade multiple hosts to reach maturity1.

K. Hoffmann & A. Chakroborty. J. Collins and A. Vieira. (Collins, Current Biology, 2017). Detailed life cycle of Schistosoma masoni.
What is trematode behavior?
Behavior, simply, is the response to a stimulus. So, trematode behavior is the response to any stimulus in the trematode’s environment. This week’s unknown organism was Schistosoma masoni and is a digenean trematode. The discussion on trematode behavior will be specifically addressing that of digenean trematodes.
Miracidia stage behavior
Miracidia is the first stage of the trematode life cycle once they hatch from their egg. During this stage they will search for their first – an unfortunate snail – of multiple hosts. The majority of mircidia are equipped with cilia to help them navigate the water column to find their host3.

D.W.T. Crompton and S.M. Joyner, 1980. Here is an example of the cilia that miracidia stage trematodes possess. Cilia is present in most species; however, there are some species that have spines instead of cilia.
Not only are miracidia good swimmers because of their cilia, they are also well adapted to finding their hosts with sensory structures called apical papilla7. These papilla contribute greatly to trematode behavior as they integrate the environmental cues that lead a miracidia to its host. In order to hone in on their potential hosts, miracidia use chemical cues the snail secretes to deduce its general location4,6. The use of a chemical stimulus to induce movement can be categorized as chemokinesis or chemotaxis.
Chemokinesis (chemo – chemical; kinesis – movement) is random movement of the organism in response to the chemical cue. Chemotaxis (chemo – chemical; taxis – movement in a determined direction) is the directional movement of the organism in response to the chemical cue.
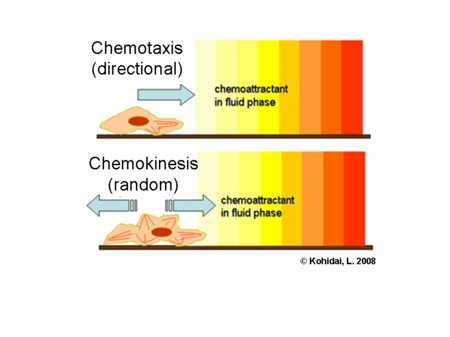
Creative Commons, WIKI. Observe the difference between the upper and lower panel. The red dot represents the organism (for our sake, a trematode miracidia stage) and their movement.
Most trematodes move randomly (chemokinesis) rather than directionally when stimulated with chemical cues from the host4,6. D.W.T. Crompton described this movement as “…a frantic, underwater devil-dance” and hypothesized that it increased the likelihood of coming in contact with the host (snail)5.
But with all the organisms thriving in a freshwater environment it seems that multiple chemical cues could disorient a trematode miracidia. ~Thankfully~ for our trematode friends, they’ve learned to specialize. Species of trematodes will only be attracted to the chemical cues of hosts with which they are compatible4.
What can they do once they’ve found their host? Miracidia will attach to the snail’s soft parts with their sucking mouth which triggers secretions to be poured onto the snail’s skin3. These secretions damage the epithelial and sub-epithelial cells; the damaged area is easily accessed by the miracidia by thrusting itself into the snail’s wound3.
While in the snail, the miracidia will develop and emerge as cercariae larvae.
Cercariae stage behavior
Compared to miracidia, cercariae are in pursuit of hosts that are more active, and thus must have a way to increase the likelihood of attaching to one. The cercariae are equipped with a tail that can propel them through the water column4.

D.W.T. Crompton and S.M. Joyner, 1980. Observe the differences in appearance of the cercariae stages. All, however, possess a very strong swimming tail.
While most contact with potential hosts is purely by chance, mixing bouts of swimming with floating, eye spots containing photoreceptors that react to changes in light stimulate a rapid swimming behavior in cercariae3,4. This excited swimming is similar to that of the miracidia. Cercariae are also sensitive to water currents and tactical feedback from brushing up against a potential host3.
It is important to note that while these stimuli (shadow, water current, and touching) could be an indicator of potential hosts, it is also likely that the cercariae makes contact with floating debris – an unsuitable host.
Once the cercariae find a suitable host, they utilize temperature gradients and chemical cues to attach to the host and find sites to penetrate the skin4. When looking for promising entry sites, the cercariae use chemical gradients in determining the vulnerability of the skin; wrinkles and hair projections have been observed as the preferred entry sites5.
When a preferable entry site is found, they anchor themselves to the host by secreting an adhesive5. By using this mechanism to ensure their stability, the cercariae are able to begin secreting chemicals that deteriorate the epithelial cells, which – with their thrusting – will gain them access to the hosts body5.
Inside the body, the cercariae navigate to the blood stream using chemical and temperature cues and eventually make their way to the liver or abdominal blood vessels to develop5.
Similarities
Recall that the phylum platyhelminthes includes the classes turbellaria, cestoda, monogenea, and trematoda. Cestoda, monogenea, and trematoda are further classified as neodermata (meaning “new skin”)1,2. Below is a phylogenetic tree representing the phylum. Only the orders for turbellaria are shown.

Laumer et al., 2015. Creative Commons. J. Sikes. (Collins, Current Biology, 2017). The phylogenetic tree shows the evolutionary relationships between the classes. Cestoda, Monogenea, and Trematoda are grouped together in a group called the Neodermata (meaning “new skin”).
All four classes possess a highly complex nervous system that helps them integrate cues from their environments; furthermore, this neural complexity helps them perform complex behaviors because of these cues2. These behaviors can include the hunting behavior of carnivorous turbellaria, the attraction between miracidia and their host, or the secretion of digestive chemicals applied to the skin of host organisms2,3,4,5.
Aside from trematodes, behavior characteristics are most well known about turbellarians. Much like the photoreceptors on trematodes (being used to detect shadows in some species), many free-living turbellarians have at least one pair of eyes1. Both classes avoid light to avoid predation.
Species of turbellarians use similar direction orientation mechanisms when finding a desired cue (whether that be food or a host). Turbellarians use trial-and-error swimming to locate prey; similarly, trematodes use chemokinesis (random movement) to find their host3.
Not much is known about the behavior of cestoda and monogenea classes. In effort to hypothesize about their behavior, cestodes must have a way of sensing their location within a host. It is possible that, like trematodes, cestodes are able to use chemical signaling to asses their surroundings. It is thought to be similar for monogeneans in the sense that they are also able to use chemical and/or temperature cues to determine when they come into contact with their hosts.
Differences
Within this phylum there are many differences. Firstly, turbellarians are free living carnivores or commensalists2. This alters the cues they receive and ultimately their behavior.
While both turbellarians and trematodes have photoreceptors, turbellarians have more complex eyes which are able to detect a bigger difference in light; similarly, trematode photoreceptors can only distinguish shadows.
Trematodes are highly sensitive to tactile stimuli and will try to attach to substrates, plants, or whatever material touches it3. In contrast, turbellarians move towards tactile stimuli touching their undersides and away from tactile stimuli touching their dorsal side8.
The class monogenea are ectoparasites, meaning they live on the outside of their host2. This means that once they find their host, cues do not have to be processed to create a navigation system to the correct location in the host body.
Many of the miracidia are ciliated, but species such as Otodistomum cestoides and Halipegus eccentricus have spines (figure 7c)3. Since they do not have cilia, their swimming capabilities are not great, and they meet their hosts by chance.

D.W.T. Crompton and S.M. Joyner, 1980. In the above figure, A and B represent ciliated miracidia, whereas C shows a miracidia with spines.
Cercariae can exhibit two forms of host entry depending on species of trematode. Passive entry into the host is by way of ingestion, enticing the primary host by using the intermediate host as bait (primary host ingests intermediate), or by passing through the digestive system of the intermediate host and attracting the primary host through the excrement3.
Trematode behavior varies widely across classes. What is similar across the phylum is that they can perform complex behaviors in response to environmental stimuli. That characteristic alone makes them an ideal candidate for behavioral research.
Works Cited
1 Pechenik, JA. Biology of the Invertebrates. 7th ed., McGraw Hill Ed., 2015.
2 Collins, J. “Platyhelminthes.” Current Biology, 27(7): 252-256. 2017
3 Erasmus, D.A. Biology of Trematodes. Edward Arnold Publishers. 1972.
4 Haas, W. “Parasitic worms: strategies of host finding, recognition, and invasion.” Zoology, 106: 349-364. 2003.
5 Crompton, D.W.T. and S.M. Joyner. Parasitic Worms. Wykeham Publications. 1980.
6 Chernin, E. “Behavioral Responses of Miracidia of Schistosome mansion and Other Trematodes to Substances Emitted by Snails.” Parasitology, 56(2): 287-296. 1970.
7 Blankenspoor, H.D., and H. van der Schalie. “Attachment and Penetrationof Miracidia Observed by Scanning Electron Microscopy.” Science, 191(4224): 291-293. 1976.
8 “Turbellarians.” 2017. http://animals.jrank.org/pages/1509/Turbellarians-Turbellaria-BEHAVIOR-REPRODUCTION.html